Pressmeddelande: Nobelpriset i kemi 1997
English
Swedish
![]()
15 October 1997
Kungl. Vetenskapsakademien har beslutat utdela 1997 års Nobelpris i kemi med ena hälften till:
Professor Paul D. Boyer, University of California, Los Angeles, USA och
Dr. John E. Walker, Medical Research Council Laboratory of Molecular Biology, Cambridge, Storbritannien
för deras klargörande av den enzymatiska mekanismen för syntes av adenosintrifosfat (ATP)
och med andra hälften till:
Professor Jens C. Skou, Aarhus Universitet, Danmark
för det först upptäckta jontransporterande enzymet, Na+, K+-ATPas
De tre pristagarna har gjort banbrytande arbeten över enzymer, som deltar i omsättningen av “högenergi” föreningen adenosintrifosfat (ATP).
Paul D. Boyer och John E. Walker får halva priset för sina arbeten över hur enzymet ATP-syntas styr bildningen av ATP. Boyer och hans medarbetare har på grundval av biokemiska data föreslagit en mekanism för hur ATP bildas ur adenosindifosfat (ADP) och oorganiskt fosfat. Walker och hans medarbetare har fastställt strukturen för enzymet och verifierat den av Boyer föreslagna mekanismen.
Jens C. Skou erhåller den andra hälften för upptäckten av enzymet natrium, kalium-stimulerat adenosintrifosfatas (Na+, K+-ATPas). Detta enzym upprätthåller balansen av natrium- och kaliumjoner i den levande cellen.
Båda enzymerna är bundna till membran i cellen och kopplade till transport av joner över dessa – dock av olika orsak.
ATP – den universella energitransportören i den levande cellen
Den tyske kemisten Karl Lohmann upptäckte ATP 1929. Strukturen klarlades några år senare och 1948 kunde 1957 års Nobelpristagare, skotten Alexander Todd, kemiskt syntetisera ATP. En betydelsefull roll spelade 1953 års medicinpristagare Fritz Lipmann när han under åren 1939-41 visade att ATP är den huvudsakliga bäraren av kemisk energi i cellen och myntade begreppet “energirika fosfatbindningar”.
ATP fungerar som energitransportör hos alla levande organismer från bakterier och svampar till växter och djur inklusive människor. ATP fångar upp den kemiska energi, som erhålls vid förbränning av födoämnen och överför den till reaktioner, som fordrar tillförsel av energi, t ex uppbyggnad av cellens beståndsdelar, muskelkontraktion, överföring av nervbudskap och mycket annat. ATP kallas cellens energivaluta.
Adenosintrifosfat (ATP) är uppbyggt av nukleosiden adenosin kopplad till tre fosfatgrupper i rad. Vid avspjälkning av den yttersta fosfatgruppen bildas adenosindifosfat (ADP) samtidigt som den frigjorda energin kan tas om hand för andra reaktioner. Omvänt kan med hjälp av energi en oorganisk fosfatgrupp fästas till ADP och bilda ATP. Det är ansenliga mängder ATP, som bildas och förbrukas. I vila omsätter en vuxen person dagligen en mängd ATP, som motsvarar ungefär halva kroppsvikten, och under hårt arbete kan mängden uppgå till nästan ett ton. Den övervägande delen av ATP-syntesen sker med hjälp av enzymet ATP-syntas. I vila förbrukar Na+, K+-ATPas en tredjedel av allt ATP som bildas.
ATP-syntaset – en särpräglad molekylär maskin
Under 1940 och 50-talen klarlades att huvuddelen av ATP bildas vid cellandningen i mitokondrierna och vid fotosyntesen i växternas kloroplaster. År 1960 isolerade amerikanen Efraim Racker och medarbetare från mitokondrier enzymet “Fo F1-ATPas” som vi nu kallar ATP-syntas. Enzymet kan uppdelas i en F1-del som innehåller den enzymatiska aktiviteten och en Fo-del som kopplar F1-delen till membranet. Samma enzym finns i kloroplaster och bakterier. År 1961 beskrev Peter Mitchell den s.k. kemiosmotiska hypotesen för vilken han 1978 fick Nobelpris. Han visade att cellandning leder till skilda koncentrationer av vätejoner (pH) innanför och utanför mitokondriemembranet och att en ström av vätejoner driver bildningen av ATP. Samma sak gäller kloroplastmembranet. Kopplingen av ATP-syntas till vätejontransport sker via Fo-delen.
Paul D. Boyer påbörjade sina studier av ATP-bildning redan på 1950-talet och är fortfarande synnerligen aktiv. Hans huvudsakliga intresse har varit att med isotopteknik ta reda på hur ATP-syntas fungerar och särskilt hur det utnyttjar energin för att bilda nytt ATP. Hans arbete har krönts med ovanlig framgång de allra senaste åren. ATP-syntas har ett för enzymer ovanligt funktionssätt, som krävt lång tid och omfattande studier att klarlägga. John E. Walker gjorde sina första studier av ATP-syntas i början av 1980-talet. Hans utgångspunkt var att det krävs en detaljerad kemisk och strukturell kunskap om ett enzym, för att förstå hur det i detalj fungerar. Han bestämde därför aminosyra-sekvenserna hos de proteinenheter som ingår. Under 1990-talet har han samverkat med kristallografer för att klarlägga den tredimensionella strukturen av ATP-syntas. Man har hitintills bestämt strukturen för enzymets F1-del. Walkers arbeten kompletterar Boyers på ett anmärkningsvärt sätt och de fortsatta studier, som baserar sig på denna struktur, har vidimerat riktigheten av den av Boyer beskrivna mekanismen.
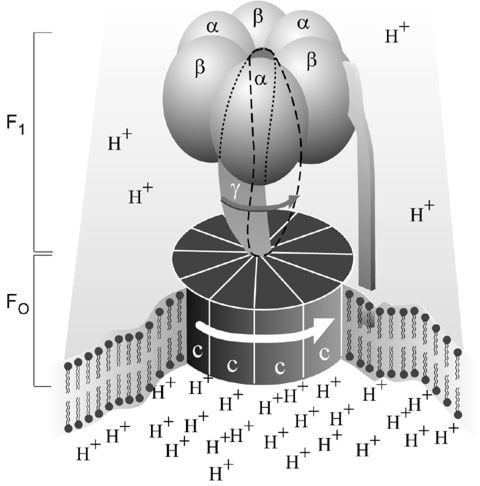
Figur 1. En förenklad bild av ATP-syntas I membranet sitter den del (Fo ) genom vilken vätejoner (H+ ) strömmar. Utanför membranet sitter den del (F1 ), som syntetiserar ATP. När vätejonerna flödar genom membranet via skivan av c-underheter i Fo-delen, tvingas skivan att vridas runt. Gamma-underenheten i F1-delen är fäst vid skivan och roterar därför med. De tre alfa- och tre beta-underenheterna i F1-delen kan däremot inte rotera. De är låsta i ett fast läge av b-underenheten. Den är i sin tur förankrad i membranet. Gamma-underenheten roterar alltså inne i den cylinder som bildas av de sex alfa- och beta-underenheterna. Genom att gamma-underenheten är asymmetrisk tvingar den beta-underenheterna till strukturförändringar. Detta leder till att beta-underenheterna binder ATP och ADP olika starkt (se Fig. 2).
ATP-syntaset (Fig. 1) består som nämnts ovan av en membranbunden del (Fo), som transporterar vätejoner, och en utskjutande del (F1) som kan frigöras från membranet. (Beteckningarna är historiska. F1 står för faktor 1 och Fo för oligomycinkänslig faktor.) Varje Fo-del består av tre typer av underenheter i olika antal, proteinerna a (1 st), b (2 st) och c (9-12 st). F1-delen består av fem olika underenheter, proteinerna alfa, beta, gamma, delta, och epsilon. Medan det finns tre av vardera alfa och beta finns det bara en enhet av var av de övriga. Det har visat sig att det är på beta-enheterna som syntesen av ATP sker. Genom den analys av aminosyra-sekvenserna som Walker och medarbetare gjorde i början av 1980-talet har det visats att underenheterna gamma, delta och epsilon inte är symmetriska, ett särdrag av betydelse för vår förståelse av hur ATP-syntaset fungerar.
De mest ingående studierna av ATP-syntaset gäller F1-delen och dess funktionssätt. Boyer och medarbetare har kunnat klarlägga att enzymet fungerar på ett mycket speciellt sätt. De fann till skillnad från den allmänna föreställningen att det energikrävande steget inte var syntesen av ATP från ADP och oorganiskt fosfat utan att det krävdes energi för att binda ADP och fosfatet till enzymet liksom för att frisätta ATP. I detta avseende skiljer sig ATP-syntaset från merparten av alla enzymer, som binder och frisätter substrat och produkter spontant, men där själva den katalytiska reaktionen kräver energi. En annan observation var, att trots den asymmetriska karaktären hos F1 finns det bara ett sätt för enzymet att reagera. Men hur kan då de tre beta-underenheterna fungera på samma sätt om de har olika kopplingar till underenheterna gamma, delta och epsilon? Boyer fann svaret på denna fråga genom att klargöra att gamma, delta och epsilon roterar i en cylinder, som bildas av alternerande alfa- och beta-underenheter. Denna rotation förorsakar strukturförändringar i beta, som leder till skillnader i bindningsförmåga för ADP och ATP under ett cykliskt förlopp (se figur 2). Denna mekanism kallas Boyers “Binding Change Mechanism”. Boyer föreslog även att denna rotation drivs av det vätejonflöde genom membranet som nämnts ovan.
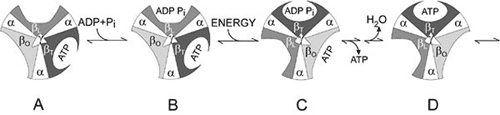
Figur 2. Boyers “Binding Change Mechanism” Bilden visar cylindern med alternerande alfa- och beta-underenheter vid fyra olika stadier av ATP-syntesen. I mitten syns den asymmetriska gamma-underenheten som orsakar förändringar i beta-underenheternas struktur. Strukturerna benämns öppen betaO (ljusgrå sektor), lös betaL (grå sektor) och tät betaT (svart sektor). I stadium A ser vi en redan färdigbildad ATP-molekyl bunden till betaT. I steget till stadium B binder betaL ADP och oorganiskt fosfat (Pi ). I nästa stadium C ser vi hur gamma-underenheten har vridit sig p.g.a. flödet av vätejoner (se fig.1). Detta leder till att de tre beta-underenheternas struktur förändrats. Den täta beta-underenheten blir nu öppen och den bundna ATP-molekylen frisätts. Den lösa beta-underenheten blir tät och den öppna blir lös. I det sista steget D sker den kemiska reaktionen då fosfatjonen reagerar med ADP-molekylen så att en ny ATP-molekyl bildas. Vi är då tillbaka till det första stadiet.
Boyer har kallat ATP-syntaset en molekylär maskin. Man kan likna det vid en vattendriven hammare, som präglar mynt. Fo-delen är vattenhjulet, flödet av protoner är vattenfallet och strukturförändringarna i F1 leder till att tre mynt av ATP-valutan hamras ut för varje varv som hjulet gör.
Walker har klarlagt de strukturella förutsättningarna för enzymets molekylära maskineri och därigenom möjliggjort en förståelse för Boyers mekanism. Den kristallografiska strukturen av F1-delen av ATP-syntas från ko, som bestämts framför allt i samarbete med holländaren J.P. Abrahams och engelsmannen A. Leslie, visar dels att alfa- och beta-underenheterna är strukturellt och evolutionärt besläktade, dels att de tre beta-underenheterna har tydligt olika struktur och därför olika förmåga att binda ADP och ATP. Gamma-underenheten är belägen som en asymmetrisk axel i den cylinder, som bildas av de 3 alfa- och de 3 beta-underenheterna och den har unika kontakter med beta-underenheterna och tvingar deras aktiva ytor att inta olika tredimensionella strukturer. Dessa resultat kan tolkas i enlighet med Boyers mekanism att enzymet fungerar genom rotation av gamma-underenheten. Det har varit svårt att experimentellt visa denna rotation men flera olika grupper har nu lyckats. Wolfgang Junge i Tyskland har använt spektroskopisk teknik och amerikanen Richard Cross har utnyttjat kemisk tvärbindning. Nyligen har en japansk grupp ledd av Masasuke Yoshida kunnat synliggöra rotationen i F1-delen av ATP-syntaset. De fäste en fiber av muskelproteinet aktin vid gamma-underenheten och beta-enheterna fästes vid underlaget. Beroende på ATP-koncentrationen i den omslutande vätskan kunde man i mikroskop visa hur aktinfibern roterade allt fortare med ökande koncentration av ATP.
Na+, K+-ATPas, den först upptäckta molekylära pumpen
Redan på 1920-talet var det känt att jonsammansättningen inuti levande celler är annorlunda än i omgivningen. Inuti cellerna är natriumkoncentrationen lägre och kaliumkoncentrationen högre än i vätskan utanför. Genom engelsmännen Richard Keynes och Alan Hodgkin (Hodgkin fick Nobelpris 1963) arbeten i början av 1950-talet blev det känt att vid en nervretning strömmar natriumjoner in i nervcellen. Skillnaden i koncentration återställs genom att natrium ånyo transporteras ut. Att denna transport fordrade ATP var sannolik eftersom man kunde hämma den i den levande cellen genom att hämma ATP-bildningen.
Med detta som utgångspunkt började Jens C. Skou söka efter ett ATP-nedbrytande enzym i nervmembran, som kunde kopplas till jontransport och 1957 publicerade han den första artikeln om ett ATPas, som aktiveras av natrium- och kaliumjoner (Na+, K+-ATPas). Han var den förste som beskrev ett enzym som kan ge en riktad (vektoriell) transport av substanser över ett cellmembran, en fundamental egenskap hos varje levande cell. Sedermera har man påvisat en mångfald enzymer med principiellt liknande verkan.
Skou använde som försöksmaterial finfördelade nervmembraner från krabba. Det ATP-spjälkade enzym, som fanns i preparatet, fordrade närvaro av magnesiumjoner och stimulerades av ökande mängder natriumjoner upp till en viss gräns. Utöver detta kunde han få en ytterligare stimulering om han tillsatte små mängder kaliumjoner. En indikation på att enzymet var kopplat till jonpumpen var att maximal stimulering erhölls vid de koncentrationer av natrium och kalium, som normalt föreligger i nerven. I sina fortsatta studier av enzymmekanismen visade Skou att natriumjoner och kaliumjoner fäster sig med hög affinitet på olika platser i enzymet. Vidare kunde han visa att den från ATP avspjälkade fosfatgruppen också fäster sig vid ATPaset. Man säger att enzymet fosforyleras. Enzymet är beroende av natriumjoner när det fosforyleras och kaliumjoner när fosfor spjälkas loss. Substanser, som är kända för att hämma natrium/kalium transport är vissa alkaloider från Digitalis (fingerborgsblomman), t. ex. oubain, och Skou visade att oubain interfererar i natriumaktiveringen av enzymet.
Den bild som så småningom växte fram genom Skous och andras arbeten är att enzymet består av två underenheter, alfa och beta. Den första bär enzymaktiviteten och den andra stabiliserar förmodligen strukturen. Enzymmolekylerna sitter i cellens yttermembran, ofta två och två, och exponerar ytor mot både ut och insidan. På den yta, som vätter mot cellens insida fäster sig tre natriumjoner och ATP. Därefter överföres en fosfatgrupp från ATP till en aminosyra i enzymet, asparaginsyra, varefter ADP lossnar och enzymet ändrar form så att natriumjonerna förs över till utsidan. Här frigörs de och i stället fäster sig två kaliumjoner på proteinet. När det fosfat som är bundet till enzymet spjälkas av förs kaliumjonerna in i cellen och när nytt ATP binder sig till enzymet avstöts de.
Resultatet av Na+, K+-ATPasets aktivitet är att cellen bibehåller en hög kaliumkoncentration. Eftersom cellmembranet är ganska genomträngligt för kaliumjoner läcker en del av dessa ut men lämnar negativt laddade joner kvar på insidan utan möjlighet att komma ut. Därför blir cellens insida negativt laddad i jämförelse med utsidan.
Spänningsskillnaden över cellmembranet är en förutsättning för att en nervretning skall kunna fortplanta sig längs en nervfiber eller en muskelcell. Därför leder en brist på näringsämnen eller syrgas i hjärnan snabbt till medvetslöshet då ATP-bindningen upphör och jonpumpen stoppar. Pumpen är också betydelsefull för att upprätthålla volymen av en cell. Stoppar pumpen sväller cellen. Skillnaden i natriumkoncentration mellan in- och utsida är drivkraft i upptag av viktiga näringsämnen, som cellen behöver t ex glukos och aminosyror. Den kan också användas för utbyte med andra joner. Sålunda kan natriumjoner, som går in, utbytas mot kalciumjoner, som går ut.
Efter upptäckten av Na+, K+-ATPas har ett antal andra jonpumpar upptäckts med likartad struktur och funktion. Som exempel kan nämnas Ca2+-ATPas i skelettmuskel, som deltar i styrningen av muskelkontraktionen, och H+, K+-ATPas, som producerar saltsyra i magsäcken. Det är det senare enzymet, som specifikt hämmas vid modern magsårsbehandling. Motsvarande enzymer finns också i lägre organismer t.ex. i jäst där ett H+-ATPas utsöndrar vätejoner, som bildas under jäsningen. Med ett samlingsnamn kallas dessa enzymer idag P-typ ATPaser eftersom de fosforyleras (engelska, phosphorylation) under reaktionens gång.
| Lästips |
| Paul D. Boyer och John E. Walker |
| Boyer, P.D., The binding change mechanism for ATP synthase – Some probabilities and possibilities, Biochimica et Biophysica Acta (1993) 1140, 215-250. |
| Abrahams, J.P., Leslie, A.G., Lutter, R., and Walker J.E., Structure at 2.8 Å resolution of F1-ATPase from bovine heart mitochondria, Nature (1994) 370, 621-628. |
| Boyer, P.D., The ATP synthase – a splendid molecular machine, Annual Review in Biochemistry (1997) 66, 717-749. |
| Jens C. Skou |
| Skou, J.C., The influence of some cations on an adenosine triphosphatase from peripheral nerves, Biochimica et Biophysica Acta (1957) 23, 394-401. |
| Skou, J.C., and Esmann, M., The Na, K-ATPase, Journal of Bioenergetics and Biomembranes (1992) 24, 249-261. |
| Lingrel, J.B., Na-K-ATPase: Isoform Structure, Function, and Expression, Journal of Bioenergetics and Biomembranes (1992) 24, 263-270. |
| Möller, J.V., Juul, B., and le Maire, M., Structural organization, ion transport, and energy transduction of P-type ATPases, Biochimica et Biophysica Acta (1996) 1286, 1-51. |
| Lutsenko, S. and Kaplan, J.H., Organization of P-type ATPases: Significance of structural diversity, Biochemistry (1996) 34, 15607-15613. |
Professor Paul D. Boyer, född 1918 i Provo, Utah, USA. Ph.D. i biokemi 1943 vid University of Wisconsin, Madison, USA. 1963-1989 var han professor i kemi vid Department of Chemistry & Biochemistry, University of California at Los Angeles (UCLA) och 1965-1983 Director vid the Molecular Biology Institute, UCLA. Sedan 1990 han är verksam som professor emeritus vid Department of Chemistry & Biochemistry, UCLA. Boyer är sedan 1970 medlem av the National Academy of Sciences, blev hedersdoktor vid Stockholms universitet 1974 och belönades 1989 av American Society of Biochemistry and Molecular Biology med the Rose Award.
Professor Paul D. Boyer
Department of Chemistry & Biochemistry
University of California
Los Angeles, CA 90024, USA
Dr. John E. Walker är född 1941 i Halifax, Storbritannien. Han erhöll M.A. och Dr. Phil. vid Oxford University, Storbritannien. Walker har sedan 1982 varit Senior Scientist vid Medical Research Council Laboratory of Molecular Biology, Cambridge, Storbritannien. Han valdes in i Royal Society, London, 1995.
Dr. John E. Walker
Medical Research Council Laboratory of Molecular Biology
Hills Road
Cambridge, CB2 2QH
Storbritannien
Professor Jens C. Skou är född 1918 i Danmark. Han fick sin medicinska utbildning vid Köpenhamns universitet. Skou disputerade 1954 vid Aarhus universitet för medicine doktorsgrad och blev där 1963 utnämnd till professor i fysiologi. 1977 utnämndes han till professor i biofysik vid samma lärosäte. Skou är ledamot av den danska vetenskapsakademien. Han har erhållit flera vetenskapliga pris bl.a. Svenska Läkaresällskapets Anders Retzius-medalj och 1985 Eric K. Fernströms stora pris.
Professor Jens C. Skou
Aarhus Universitet
Nordre Ringgade
DK-8000 Aarhus
Danmark
Nobel Prizes and laureates
Six prizes were awarded for achievements that have conferred the greatest benefit to humankind. The 14 laureates' work and discoveries range from quantum tunnelling to promoting democratic rights.
See them all presented here.

